
ОНТОГЕНЕЗ ВЫСШИХ РАСТЕНИЙ.
Онтогенезом называют индивидуальное развитие организма от зиготы или вегетативного зачатка до естественной смерти. В ходе онтогенеза реализуется наследственная информация организма – его генотип– в конкретных условиях окружающей среды, в ре-зультате чего формируется фенотип, то есть совокупность всех признаков и свойств данного индивидуального организма. Онтогенез включает в себя все жизненные процессы и продолжается у разных растений от 10—14 дней до 3—5 тыс. лет. По продолжительности жизни растения делят на эфемеры, однолетние, двулетние и многолетние (секвойи — до 5 тыс. лет).
Независимо от продолжительности жизни все растения можно разделить на две группы: моно- и поликарпические. Монокарпическими (греч. mono — один, karpos — плод) называются растения, цветущие и плодоносящие один раз в своей жизни (эфемеры, однолетние растения, двулетние растения (мокровь, свекла) и некоторые многолетние, например мексиканские агавы, бамбук) Наступление плодоношения у таких организмов приводит к их быстрому старению и отмиранию. Поликарпические растения — это растения, плодоносящие много раз в жизни.
Онтогенез дискретен, т. е. его можно разделить на отдельные этапы, проходящие последовательно один за другим. В онтогенезе цветковых растений выделяют 5 этапов: эмбриональный, ювенильный (молодости), зрелости, размножения (генеративный) и старости (сенильный).
1.Эмбриональный этап — это период образования зародыша и семени, который начинается со слияния яйцеклетки и спермия и образования зиготы на материнском растении. Зародыш состоит из меристематических тканей и питается гетеротрофно, т. е. за счет питательных веществ, поступающих из материнского растения. У цветковых растений зигота делится поперек, образуя мате-ринскую клетку суспензора (нижняя клетка) и будущий проэмбрио (верхняя клетка). В суспензорной части образуется подвесок, продвигающий зародыш вглубь эндосперма. На стадии глобулы зародыш имеет шаровидную форму. Далее у двудольных синхрон-но растут две семядоли, что приводит к стадии сердечка, а затем торпедо. У некоторых растений на эмбриональной стадии форми-руются уникальные органы (например, щиток, колеоптиль или гаустория). Эмбриональный этап заканчивается полным формиро-ванием семян и переходом их в состояние покоя.
2. Ювенильный этап (или молодость) — это период от прорастания семени до начала заложения первых цветков. Прорастание семени происходит при наступлении благоприятных условий после периода покоя. Оно является возобновлением роста в резуль-тате поступления в семя воды и его набухания. Содержащиеся в семени ферменты активизируются, а также синтезируются новые ферменты. В процессе своего формирования проросток постепенно переходит с гетеротрофного на автотрофное питание. Для ювенильных растений характерна максимальная активность всех физиологических функций, поэтому растения в это время обла-дают минимальной устойчивостью. У молодых растений большая способность к корнеобразованию: черенки, срезанные в этот период онтогенеза, легко укореняются, что с давних пор используется в садоводстве и лесоводстве. На этом этапе у растения обра-зуются только вегетативные органы: листья, стебли, корни.
3. Зрелость — период формирования репродуктивных органов растения от заложения первого цветка до первого оплодотворения. Заложение цветков тормозит рост вегетативных органов. В определенный этап онтогенеза верхушечная меристема побега начи-нает вместо листьев, междоузлий формировать цветки или соцветия. Однако она должна быть индуцирована для перехода к генеративному развитию. Внешними индукторами этого процесса являются температура, свет, продолжительность дня и ночи, вода, элементы минерального питания.
У некоторых растений способность к заложению цветков, т. е. переход к этапу зрелости, появляется лишь после действия на них пониженных температур в течение определенного времени. Уже в старину люди знали, что злаки делятся на двулетние и однолет-ние. Однолетние злаки колосятся в первый год и называются яровыми, а двулетние — только после перезимовки и называются озимыми. Свойство озимых однолетних и двулетних растений ускорять переход к заложению цветков после действия на них пони-женных температур в течение определенного времени назвали яровизацией. Продолжительность периода охлаждения и эффек-тивные температуры зависят от вида и даже разновидности растений. У большинства растений этот период составляет 1—3 мес, у других— от нескольких дней до двух недель. Для сельдерея, хризантемы, плевела многолетнего, гравилата и левкоя достаточно 1 —2-дневного охлаждения.
В зависимости от реакции на длину дня, растения делятся на короткодневные, переходящие к цветению только тогда, когда день короче ночи (день составляет 8—12 ч в сут) — рис, кукуруза, просо, соя, сахарный тростник, хлопчатник, сорго; длиннодневные ( день не менее 16—18 ч/сут) — пщеница, ячмень, овес, горчица, свекла, лен, шпинат, клевер,укроп; растения, нуждающиеся в чере-довании разных фотопериодов, а также нейтральные по отношению к длине дня (гречиха, горох, фасоль, томаты).
Продолжительность дня или ночи (фотопериод) воспринимает листовая пластинка. Основную роль в восприятии фотопериода играет фитохром, а изменения, в результате которых начинается заложение цветков, происходят в меристеме — ближайшей к листу точке роста. Длинно- или короткодневность растений зависит от географического происхождения вида или сорта. Длинноднев-ность выработалась у растений в связи с перезимовкой, короткодневность — в связи с периодическими засухами или тропически-ми ливнями. Длиннодневные растения распространены, в основном, в умеренных и приполярных широтах, короткодневные – в субтропиках.
В листьях, получающих необходимый фотопериод, синтезируются вещества, которые поступают в верхушечные меристемы и вызывают цветение. М.Х. Чайлахян выдвинуть гипотезу, согласно которой цветение вызывают два гормона: один из них — гиббе-реллин, а другой — неизвестный гормон, названный антезином. Процесс зацветания проходит в две фазы: на первой фазе обра-зуется цветочный стебель (цветоножка или ось соцветия и цветоножки), а на второй — сами цветки.
У длиннодневных растений критической является первая фаза зацветания, которая зависит от присутствия гиббереллина, синте-зирующегося в листьях на длинном дне. Антезин у этих растений всегда имеется в достаточном количестве.
Короткодневные растения, наоборот, при любой продолжительности дня содержат много гиббереллинов, в листьях при благопри-ятной продолжительности дня образуется недостающий гормон антезин.
Нейтральные растения цветут при любой длине дня, так как у них есть и гиббереллины, и антезины; образование цветков у них зависит только от возраста растения.
Формирование мужских и женских цветков
У растений чаще образуются обоеполые (гермафродитные) цветки, реже однополые — мужские или женские. Регуляция пола у раздельнополых растений контролируется фитохромом.
Короткий день или уменьшение интенсивности света вызывает массовое появление на мужских растениях обоеполых или даже женских цветков. Длинный день действует в противоположном направлении. Синий свет усиливает женскую сексуализацию, а красный — мужскую.
Однако в системе, регулирующей сексуализацию цветков, есть главные факторы -гормоны: цитокинины и гиббереллины. В женских цветках цитокининов содержится значительно больше, чем в мужских. Цитокинины, образующиеся в корнях, транспортируются в верхушки стеблей и индуцируют образование в меристеме женских (пестичных) цветков. Гиббереллины, синтезируемые в листьях, транспортируясь в верхушечные меристемы, вызывают образование мужских (тычиночных) цветков.
На заложение мужских или женских цветков влияют условия минерального питания. Установлено, что избыток солей кальция в почве способствует развитию большого количества мужских цветков, а внесение фосфорных удобрений, напротив, приводит к уве-личению числа женских цветков.
Повышенная влажность почвы и воздуха в период заложения цветков — необходимое условие для образования женских цветков, а понижение влажности способствует формированию мужских цветков. Окись углерода, этилен, ацетилен стимулируют образование женских цветков. Низкие температуры способствуют формированию женских цветков, а высокие — мужских.

Короткий день, синий свет, СО2, высокий уровень азота, высокая влажность и низкие температуры, способствующие корнеобразованию, усиливают формирование женских цветков. Длинный день, красный свет, высокий уровень кальция, высокие температуры и относительно низкая влажность, вызывающие усиление роста побегов, индуцируют образо-вание мужских цветков.
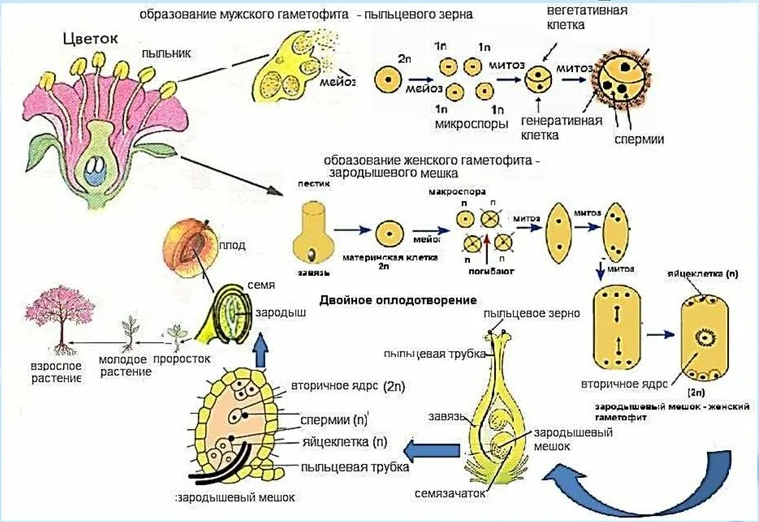
4. Этап размножения — период от первого оплодотворения до полного созревания плодов. Значение полового процесса в фило-генезе состоит в том, что при оплодотворении образуются организмы с двойной наследственностью (материнской и отцовской), а это обеспечивает им большую устойчивость и приспособляемость к постоянно изменяющимся условиям. На этом этапе происхо-дят образование, рост и созревание плодов и семян.
Оплодотворение делят на три фазы: а) опыление, б) прорастание пыльцы и рост пыльцевой трубки в тканях пестика, в) собственно оплодотворение, то есть образование зиготы. Зигота образуется при слиянии спермия пыльцевой трубки (мужской гаметофит) с яйцеклеткой зародышевого мешка (женский гаметофит). В зародышевом мешке происходит двойное оплодотворение, так как вто-рой спермий соединяется с вторичным диплоидным ядром центральной клетки зародышевого мешка. Зародыши проходят ряд последовательных фаз развития. На последнем этапе созревания семена теряют значительное количество воды и переходят в состояние покоя, когда в тканях уменьшается содержание стимуляторов роста и увеличивается количество ингибитора роста абсцизовой кислоты.
Плод развивается из завязи цветка и, как правило, содержит семена. Плоды могут формироваться без оплодотворения и образова-ния семян. Это явление называют партенокарпией. Образование партенокарпических (бессемянных) плодов может происходить при обработке растений ауксинами и гиббереллинами. Однако обычно цветки без опыления и оплодотворения опадают.
Для ускорения созревания можно использовать ряд приемов: 1) снятие плодов с деревьев, например авокадо; 2) выдерживание плодов в атмосфере этилена; 3) обеспечение растений некоторыми метаболитами, например малатом, ускоряет созревание яблок, так как малат, являясь дыхательным субстратом, стимулирует климактерический подъем интенсивности дыхания; 4) проведение дефолиации растений — искусственного удаления листьев; 5) обработка растений десикантами — веществами, ускоряющими потерю воды созревающими растениями, в результате чего метаболиты быстрее оттекают в семена.
Для замедления созревания рекомендуется хранить плоды либо при низком содержании кислорода, либо в атмосфере азота или углекислого газа в полиэтиленовых мешках
5. Старость — период от полного прекращения плодоношения до отмирания всех вегетативных органов и смерти всего организма. Этот этап характеризуется прогрессирующим старением организма. Старение — это усиливающееся с возрастом ослабление жиз-недеятельности, которое выражается в прогрессирующем нарушении синтеза белков, ослаблении регуляции, уменьшении ско-рости физиологических процессов, увеличении количества мертвых клеток; распад веществ идет быстрее их синтеза.
На организменномуровне старение характеризуется уменьшением скорости всех физиологических процессов и, прежде всего, интенсивности фотосинтеза и дыхания, оводненности тканей, уменьшением жизнедеятельности корней, снижением скорости даль-него транспорта, ослаблением регуляции. Старение — это торможение роста, связанное с пониженным синтезом ауксина и повы-шением концентрации этилена и АБК. У старых деревьев нарушается апикальное доминирование и образуется поросль.
Выделяет несколько типов старения. Однолетние растения погибают целиком. У многолетних трав ежегодно полностью отмирает надземная часть, а корневая система остается жизнеспособной. У многих растений стареют и опадают ранее образовавшиеся листья. У листопадных деревьев осенью одновременно стареют и опадают все листья. Перед опадением листа или плода в основа-нии черешка листа или плодоножки образуется отделительный слой, где размягчаются и частично растворяются клеточные стенки и срединные пластинки. Этот процесс индуцируется этиленом, продуцируемым стареющими листьями и созревающими плодами.

РАЗМНОЖЕНИЕ ВОДОРОСЛЕЙ.
У водорослей встречается бесполое и половое размножение.
Бесполое вегетативное размножение может осуществляться простым разделением многоклеточного организма на части (фраг-ментацией) или с помощью выводковых веточек или клубеньков. Бесполое размножение водорослей осуществляется также при помощи спор.

Споры водорослей образуются внутри особых клеток. При этом клетка-спорангий многократно делится на части внутри оболочки, которая затем разрывается, и продукты деления выходят наружу. Особь, на которой формируются спорангии, называют спорофи-том. Споры водорослей могут быть подвижными (зооспоры) и неподвижными (апланоспоры).
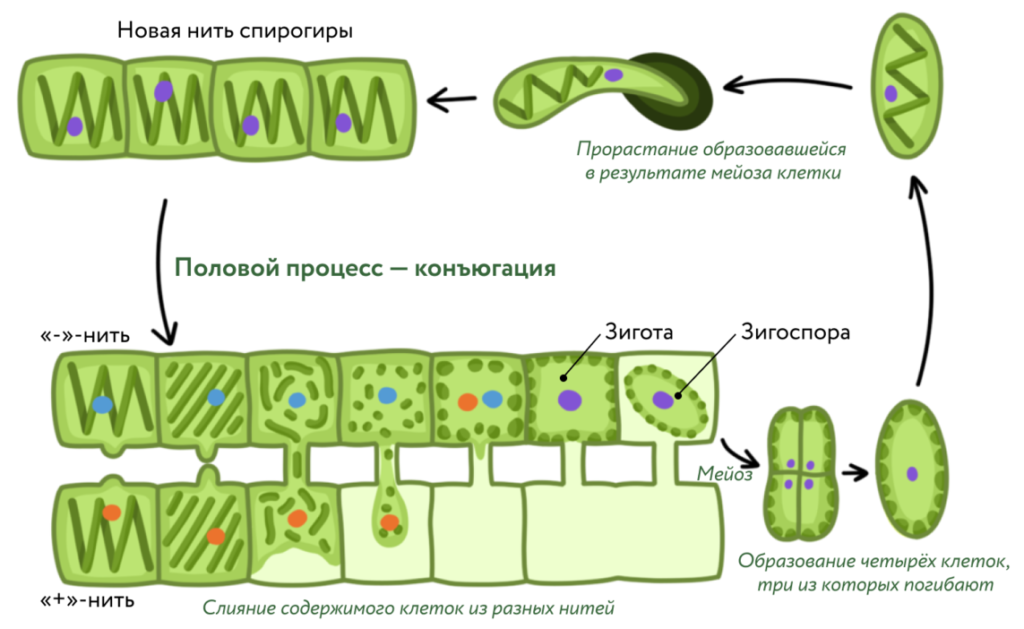
При половом размножении в клетках образуются специализированные половые клетки — гаметы, которые выходят наружу, где попарно сливаются, образуя зиготу . У некоторых водорослей специализированных половых клеток нет: образование зиготы происходит путём слияния содержимого двух вегетативных клеток. Такой половой процесс называют конъюгацией.
Клетки, в которых образуются гаметы, называются гаметангиями, а растения, несущие гаметангии, — гаметофитами.
У водорослей встречается несколько вариантов полового процесса:
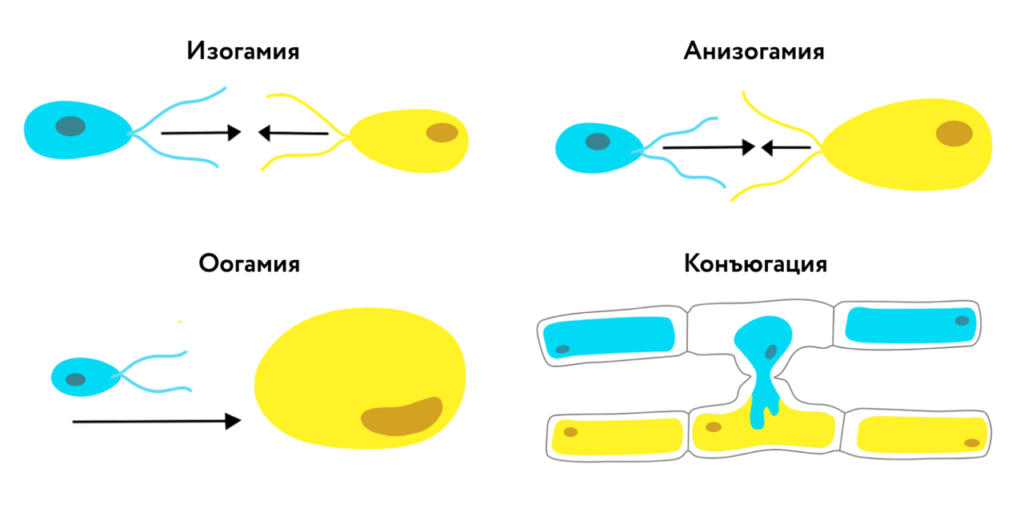
- изогамия — слияние двух одинаковых по форме и размеру подвижных гамет;
- гетерогамия — слияние двух одинаковых по форме, но разных по размеру подвижных гамет;
- оогамия (овогамия) — слияние крупной неподвижной яйцеклетки с мелким подвижным сперматозоидом;
- конъюгация — слияние содержимого двух вегетативных клеток.
Образовавшаяся зигота часто покрывается плотной оболочкой и превращается в зигоспору (зигоцисту). В таком виде она пережи-дает неблагоприятные условия, например засушливый или зимний период. При наступлении благоприятных условий зигоспора либо сразу прорастает, либо сначала в ней происходит мейоз. При мейозе из неё выходят четыре или одна спора, в этом случае ещё три клетки, образовавшиеся при делении, гибнут. У разных видов споры могут быть со жгутиками (подвижными) или безжгу-тиковыми (неподвижными). Споры дают начало новым организмам.
У бурых водорослей, например у ламинарии, в жизненном цикле чередуются два поколения — бесполое (спорофит) и половое (гаметофит). Спорофит развивается из зиготы, без её деления, он диплоидный. В особых органах спорофита — спорангиях — происходит мейоз и образуются гаплоидные споры. Из этих спор вырастают гаметофиты, гаплоидное половое поколение. На гаметофитах развиваются гаметангии, в которых образуются гаметы. Во внешней среде (в воде) или прямо на женском гаметофите происходит копуляция — встреча гамет — и образуется зигота.
Гаметофиты и спорофиты водорослей могут внешне сильно различаться или не иметь выраженных морфологических отличий. Также гаметофиты могут быть однодомными (обоеполыми), когда на одной особи развиваются и мужские, и женские гаметы, или двудомными (раздельнополыми), когда мужские гаметы образуются на одних особях, а женские — на других.

РАЗМНОЖЕНИЕ ВЫСШИХ СПОРОВЫХ РАСТЕНИЙ.
У мхов, папоротников, плаунов и хвощей размножение происходит спорами. У мхов взрослым растением является гаметофит (n) — половое поколение, которое развивается при прорастании споры.

Это листостебельное растение, на побегах которого развиваются органы полового размножения — антеридии (мужские органы) и архегонии (женские органы). В антеридиях в процессе митоза образуются сперматозоиды, а в архегониях — яйцеклетка (одна). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку, при этом образуется зигота (2n). Из зиготы на гаметофите развивается спорофит (2n) в виде коробочки на ножке — спорогона. Спорофит не способен к самостоятельному су-ществованию и питается за счет питательных веществ гаметофита. В спорангиях (коробочке) в результате мейоза образуются спо-ры. Споры после созревания высыпаются и во влажной среде прорастают, давая начало новым гаметофитам.
Схема жизненного цикла мхов

У папоротников, плаунов и хвощей взрослым растением является спорофит, на котором в специальных органах — спорангиях в результате мейоза образуются споры (n). Споры после созревания высыпаются и прорастают.

При прорастании споры развивается половое поколение — гаметофит, который представлен небольшим заростком и существует очень недолго. На заростке развиваются органы полового размножения — антеридии и архегонии, в которых в результате митоза развиваются гаметы. В антеридиях образуются сперматозоиды, а в архегониях созревает яйцеклетка. Схема жизненного цикла папоротников

При наличии капельно-жидкой влаги сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку, при этом образуется зигота. Из зиготы развивается зародыш, а далее молодое растение — спорофит.

0 Комментариев