
ПРОЦЕССЫ МАТРИЧНОГО СИНТЕЗА.
Матричные процессы — это синтез биополимеров на основе матрицы.
Это одна из интересных проблем молекулярной биологии, где много еще таких механизмов нерасшифрованно. В живом организ-ме постоянно происходит наряду с распадом синтез белка. Метод линейных атомов позволил установить, что в состав клеток вхо-дит большое количество разнообразных белков и скорость синтеза их различны. Белки эритроцитов обмениваются в течении 2-3 месяцев, в тоже время белки уже обмениваются очень быстро, установлено, что основные белки нервной ткани обмениваются в течение 21 дня.
Белки в клетках органов и тканей вступают во взаимодействие с различными компонентами и поэтому в клетках должен существо-вать механизм, который бы обеспечивал безошибочный синтез белковых веществ. Это имеет значение для метаболических процессов.
Репликация ДНК.
Поскольку ДНК является молекулой наследственности, то для реализации этого качества она должна точно копировать саму себя и таким образом сохранять всю имеющуюся в исходной молекуле ДНК информацию в виде определенной последовательности нук-леотидов. Это обеспечивается за счет особого процесса, предшествующего делению любой клетки организма, который называется репликацией ДНК.
Репликация ДНК — удвоение, происходит в синтетическую (S) стадию интерфазы перед каждым делением клетки. Синтетический-S период, продолжительность несколько часов (6-10), характеризуется процессами: репликация ДНК, синтез РНК белков. Каждая хро-мосома превращается в 2 хроматиды. Набор хромосом 2п4с.
Репликация сложный процесс идущий с участием фермента (ДНК полимераз) Для репликации нужно сначала расплес-ти двойную спираль ДНК. Это делают специальные ферменты топоизомеразы, а ферменты хеликазы расщепляют водородные связи между азотистыми основаниям. Расплетенные участки ДНК очень чувствительны к повреждающим факторам. Чтобы они оставались как можно меньше времени в таком состоянии, синтез на обеих цепочках идет одно-временно. В материнской ДНК две цепи двойной спирали антипараллельны-напротив 3ʹ конца одной цепи распола-гается 5ʹ конец — другой, а фермент ДНК полимераза может перемещаться только в одном направлении — от 3ʹ конца к 5ʹ концу матричной цепи. Поэтому репликация одной половины материнской молекулы начинающийся 3ʹ нуклеоти-дом включается после расплетения двойной спирали и идет непрерывно. Репликация второй половины молекулы начинается чуть позже, и не сначала где располагается 5ʹ нуклеотид препятсвующий репликации, а на некотором расстоянии от него. ДНК полимераза при этом движется в обратную сторону, синтезируя короткий фрагмент – фраг-мент Оказаки (1000-2000 нуклеотидов). Для начала репликации ДНК коротких фрагментов этой нити требуется синтез фрагментов РНК как затравок, для чего используется особый фермент — РНК-полимераза (праймаза). Впоследствии праймеры РНК удаляются, в образовавшиеся бреши встраивается ДНК с помощью ДНК полимеразы I. Таким образом, каждая цепь ДНК используется как матрица или шаблон для построения комплементарной цепи и репликация ДНК является полуконсервативной (т.е. одна нить в новой молекуле ДНК — «старая», а вторая — новая). Отдельные фраг-менты Оказаки сшиваются ферментом ДНК лигазой.
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более рас-пространена двунаправленная репликация. Через некоторое время после начала репликации в электронный мик-роскоп можно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окружённый более протяжёнными участками нереплицированной ДНК.
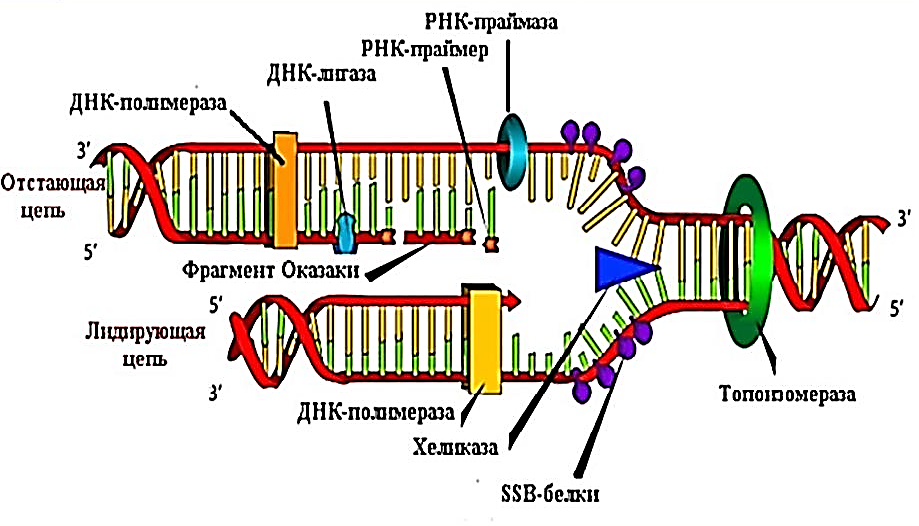
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500-5000 — у эукариот.
Скорость репликации ДНК у человека относительно низкая и для того, чтобы обеспечить репликацию ДНК любой хромосомы человека, требовались бы недели, если бы репликация начиналась из одной точки. На самом деле в молекуле ДНК любой хромосомы, а каждая хромосома человека содержит только одну молекулу ДНК, имеется мно-жество мест инициации репликации (репликонов). От каждого репликона репликация идет в обоих направлениях до тех пор, пока соседние репликоны не сливаются. Поэтому репликация ДНК в каждой хромосоме протекает относи-тельно быстро.
Репликация проходит в три этапа:
1. Инициация репликации 2. Элонгация 3. Терминация репликации.
Регуляция репликации осуществляется на этапе инициации. Репликация начинается не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как один, так и много. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хро-мосомы) состоят из большого числа самостоятельных репликонов, что сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий.
В 1957 г. Дельбрук и Стент сформулировали три альтернативные гипотезы репликации ДНК в клетках эукариот:
- консервативная репликация -исходная двухцепочечная молекула ДНК, служит матрицей для образования совершенно новой двухцепочечной молекулы, на цело достраивающейся на исходной. Одна из дочерних клеток получит исходную ДНК, а другая вновь синтезированную.
- полуконсервативная репликация-две нити ДНК расплетаются (как застёжка-молния), Каждая цепь служит матрицей для образования новой. ДНК постепенно разделяется специальным ферментом на 2 половины в продольном направлении. Каждая половинная спираль снова становится целой и вместо 1 молекулы получается две. Хромосома становится двухроматидной.
- дисперсионная репликация — исходная ДНК распадается на короткие разной длины фрагменты, используемые в качестве матриц для построения фрагментов 2-х новых двойных спиралей, которые затем воссоздаются в единую структуру молекулы. Образованные молекулы ДНК содержат старые и новые фрагменты.

Процесс репликации ДНК весьма сложен, но протекает аналогично у про- и эукариот, отличаясь участвующими ферментами, скоростью и направлением репликации, количеством точек репликации.
У прокариот репликация протекает быстрее: примерно 1000 нуклеотидов в секунду. В то время как у эукариот только около 100 нуклеотидов. Количество нуклеотидов в каждом фрагменте Оказаки у эукариот составляет примерно до 200, у прокариот — до 2000.
У прокариот кольцевые молекулы ДНК представляют собой один репликон. У эукариот каждая хромосома может содержать множество репликонов. Поэтому синтез начинается в нескольких точках, одновременно или нет.
В 1955 г. Корнберг и его коллеги открыли фермент, который обеспечивает репликацию ДНК, и назвали полимеразой. На современном этапе среди ферментов, участвующих в синтезе ДНК, выделены:
- ДНК-полимераза 1.2.3. обладают 5′->3′ полимеразной активностью.
- Топоизомеразы — ферменты, катализирующие переходы в молекулах ДНК, связанные с изменением степени сверхспирализации.
- ДНК-гиразы — переводят двухцепочечную ДНК в состояние отрицательной сверхспирализации.
Значение изучения репликации ДНК. В начале 60-х американский ученый Леонард Хейфлик обнаружил, что при культивировании в питательной среде вне организма нормальные диплоидные(соматические) клетки человека способны делиться лишь ограниченное число раз. Предельное число делений зависело от возраста того, кому принадлежали клетки, взятые в культуру .Так, клетки от новорожденных детей могли пройти 80-90 делений , в то время как клетки от 70-летних стариков делились только 20-30 раз. Максимальное число клеточных делений было названо «лимитом Хейфилка». В 1971г. отечественный ученый А.М.Оловников в своей «теории маргинотомии» предположил, что в основе ограниченного потенциала удвоения нормальных соматических клеток, растущих в культуре вне организма, может лежать постепенное укорочение ДНК хромосом с каждым раундом репликации. Хромо-сомы соматических клеток человека несут на каждом конце многократно повторенные гексамеры-TTAGGG, общая длина которых может достигать 10 тысяч пар нуклеотидов. В комплексе со специфическими белками такие тандемные повторы образуют концевые районы хромосом-теломеры. Эти специализированные структуры защищают кодирующую часть ДНК от экзонуклеаз, предотвращают неправильную рекомбинацию хромосом и позволяют им прикрепляться к ядерной оболочке. В ходе культивирования in vitro некоторых клонов нормальных клеток теломеры укорачиваются в среднем на 50 пар нуклеотидов за каждый цикл деления.
Репарация ДНК.
Процесс восстановления исходной нативной структуры ДНК называют репарацией ДНК, или генетической репарацией, а системы, участвующие в нем – репарационными. Репарация ДНК — один из важнейших генетических процессов в клетке, обеспечивающих её жизнедеятельность и сохранение вида в целом. Длина молекулы ДНК у человека составляет более 3 млрд. нуклеотидов. Нарушения в первичной структуре ДНК могут быть обусловлены: — ошибками спаривания (основание в матричной цепи ДНК в течение короткого времени может находиться в другой таутомерной форме, позволяющей присоединить в комплементарной цепи неверное основание: наиболее частая ошибка такого типа — встраивание аденина вместо цитозина с образованием пары AG, . — спонтанным отщеплением основания от цепи ДНК (депуринизация — отщепление пуринов). — дезаминированием цитозина ( превращением его в урацил) . — присоединением метильных или этильных групп к основаниям (что приводит к изменению свойств основа-ния и к образованию неверной пары). Поврежденная ДНК может индуцироваться внешними воздействиями: ультрафиолетом, рентгеновскими лучами, химическими соединениями и т.д. Ультрафиолетовое облучение (УФО) вызывает сшивку соседних тиминовых оснований в цепи ДНК. Образующиеся тиминовые димеры препятствуют нормальной репликации. Воздействие рентгеновского излучения, может вызывать одноцепочные разрывы. Более жесткое излучение приводит к образо-ванию двуцепочечных разрывов ДНК.
Классификация репарации ДНК.
- По отношению к процессу репликации различают:
А) дорепликативную репарацию — протекает в G1 периоде клеточного цикла ( фотореактивационная репарация, экс-цизионная репарация) Б) пострепликативную репарацию (осуществляется с помощью механизмов, участвующих в процессах рекомбинации и репликации ДНК).
2. По характеру протекающих процессов.
А) Фотореактивация (в 1949 г. А. Кельнер и в 1950 г. Р. Дульбекко установили, что жизнеспособность актиномицетов и бактерий, подвергнутых УФО в летальных дозах, восстанавливается, если затем воздействовать на них видимым светом. Явление было наз-вано фотореактивацией).
Б) Эксцизионная репарация ДНК. Система генетической репарации, при действии которых поврежденные участки вырезаются из цепи ДНК, отсюда происходит и термин «эксцизионная репарация» (анг. Excision — вырезание). Общая схема эксцизионной репара-ции, включает несколько этапов: 1. Узнавание повреждения УФ-эндонуклеазой 2. Инцизия (надрезание) цепи ДНК этим ферментом по обе стороны от повреждения; 3. Эксцизия (вырезание и удаление) фрагмента ДНК, содержащего повреждение, происходит при участии геликазы — фермента, расплетающего молекулу ДНК для высвобождения концов после первичных надрезов; 4. Ресинтез, в ходе которого ДНК-полимераза заполняет образовавшийся дефект в ДНК благодаря своей 5-3-полимеразной активности. Другими словами – ДНК-полимераза проводит синтез недостающего участка ДНК в соответствии с принципом комплементарности. 5. ДНК-лигаза ковалентно присоединяет вновь синтезированный участок ДНК к ранее синтезированной ДНК. В целом, эксцизионная репа-рация обычно распознает нарушения вторичной структуры ДНК (двойной спирали) и ликвидирует их.

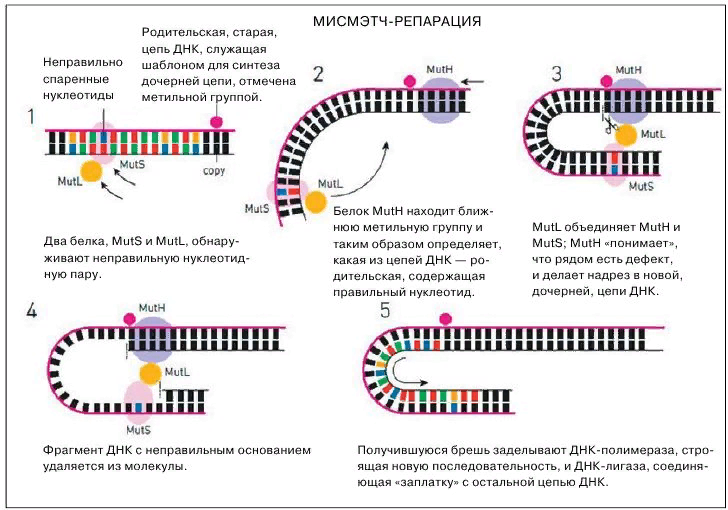
В) Исправление ошибок спаривания (мисмэтч-репарация). Мисмэтч-репарация исправляет ошибки, возникающие в результате нарушения комплементарности пар А-Т или Г-Ц в дочерней цепи при включении в них некомплементар-ных нуклеотидов. Особенность данного механизма, состоит в том, что он способен отличить «старую» цепь ДНК от «новой» и исправить именно вновь син-тезированную. В основе данного феномена лежит то важное свойство, что материнская цепь несёт в последовательностях GATC аденины с присоединенными к ним сразу после окончания репликации метильными группами. Вследствии этого во время следую-щего цикла репликации материнская и дочерняя цепи становятся структурно различными, так как до окончания данного цикла дочерняя цепь остаётся неметилированной. Именно в этот временной промежуток и должны быть исправлены ошибки спаривания осно-ваний. Генетическая репарация неспаренных оснований обнаружена в клетках и человека, и дрожжей. Этапы процесса: — к паре некомплементарных оснований прикрепляется белок MutS, затем к этому комплексу присоеди-няется белок МutL. — сформированный трехкомпонентный комплекс активирует белок mutH (который находится в клетке в неактивном, латентном состоянии) и происходит связывание с ближайшей неметилированной последова-тельностью GATC. — активный mutH надрезает дочернюю цепь ДНК около аденина, активируется другая эндонуклеаза. В результате удаляется фрагмент новой цепи, содержащий мисмэтч. — ресинтез, в ходе которого ДНК-полимераза заполняет образовавшийся дефект в ДНК благодаря своей 5-3-полимеразной актив-ности. Другими словами – ДНК-полимераза проводит синтез недостающего участка ДНК. — ДНК-лигаза ковалентно присоединяет вновь синтезированный участок ДНК к ранее синтезированной ДНК.
Г) SOS-Репарация. Системы генетической репарации, при которой точность синтеза невысока. Эти механизмы являются индуци-бельными, и обусловлены необходимостью синтеза ДНК даже на матрице, содержащей повреждения. При этом синтез ДНК на матрице, оставшейся неповрежденной, будет сопровождаться большим количеством ошибок. Хотя такая ДНК и содержит значи-тельное количество ошибок, поврежденные клетки действительно «спасаются» на каком-то этапе, если только жизненно важные функции не оказались безнадежно нарушенными. В связи со спасательными функциями этой системы репарации ДНК она была названа SOS-репарацией. Особенность прокариотических и эукариотических клеток состоит в их способности увеличивать эффек-тивность генетической репарации при высокой дозе повреждений.
Стимулом к запуску механизмов SOS-репарации служит накопление одноцепочечных разрывов ДНК. Болезни, связанные с нару-шением репарации ДНК. Пигментная ксеродерма Синдром Коккейна Трихотиодистрофия Атаксия-телеангиэктазия (синдром Луи-Бар) Анемия Фанкони Синдром Хатчинсона-Гилфорда (прогерия!) Синдром Вернера (прогерия!)
Биосинтез белка.
Транскрипция (от лат. transcriptio) — переписывание.
Все процессы метаболизма в клетке и в целом организме протекают под контролем наследственного аппарата. Рассмотрим один из важнейших процессов проявления наследственной информации в ходе пластического обмена — биосинтез белка. Он осуществляется во всех клетках про- и эукариотических организмов, это одно из свойств живого организма. Многообразие белков и их свойств определяется первичной структурой белков. т.е. последовательностью аминокислот. Информация о первичной структуре белковой молекулы закодирована последовательностью нуклеоти-дов в участке молекулы ДНК — гене. ДНК находится в ядре, а рибосомы, органоиды, на которых происходит сборка бел-ка, располагаются в цитоплазме. Информация о последовательности аминокислот и его первичной структуре должна быть доставлена к рибосоме. Для этого существует посредник — и-РНК (информационная). Этот процесс включает два этапа: транскрипцию и трансляцию.
Процесс транскрипции требует больших затрат энергии в виде АТФ и осуществляется ферментами, катализирующими процесс транскрипции: ДНК-зависимые РНК-полимеразы. У прокариот обнаружен лишь один тип этого фермента, который синтезирует все три типа РНК (мРНК, тРНК, рРНК). Эукариоты имеют три разные ДНК-зависимые РНК-полимеразы, каждая из которых ответственна за транскрип-цию генов, кодирующих разные типы клеточных РНК. Переписывание информации происходит путём синтеза на одной из цепей молекулы ДНК одноцепочечной молекулы РНК, Последовательность нуклеотидов и-РНК комплементарна (соот-ветствует) последовательности нуклеотидов матрицы — полинуклеотидной цепи ДНК.
Одновременно транскрибируется не вся молекула ДНК, а лишь отдельные её отрезки. Такой отрезок (транскриптон) начинается про-мотором — участком ДНК, куда присоединяется РНК-полимераза и откуда начинается транскрипция, а заканчивается терми-натором — участком ДНК, содержащим сигнал окончания транскрипции.
В транскрипции выделяют 3 стадии: инициация, элонгация, терминация.
Инициация транскрипции позволяет начаться синтезу молекулы РНК. Инициация включает присоединение к промотору комплекса ферментов. Главным из них является РНК-полимераза (в данном случае ДНК-зависимая), которая, в свою очередь, состоит из нес-кольких белков-субъединиц и играет роль катализатора процесса. У эукариот на инициацию транскрипции влияют особые участки ДНК: энхан-серы (усиливают) и сайленсеры (подавляют), которые обычно удалены на некоторое расстояние от самого гена. Суще-ствуют различные белковые факторы, влияющие на возможность инициации транскрипции. РНК-полимераза способна распозна-вать определенные пос-ледовательности нуклеотидов и прикрепляется к ним. Эти последовательности короткие и универсальные для всего живого.
После того, как РНК-полимераза присоединяется к промотору, участок двойной спирали ДНК раскручивается и между цепочками этого участка разрываются нуклеотидные связи. Расплетается примерно 18 пар нуклеотидов.
На стадии элонгации происходит последовательное присоединение по принципу комплиментарности свободных нуклеотидов к осво-божденному участку ДНК. РНК-полимераза соединяет нуклеотиды в полирибонуклеотидную цепочку.
При синтезе РНК около 12 ее нуклеотидов комплементарно временно связаны с нуклеотидами ДНК. При движении РНК-полимеразы впереди нее цепочки ДНК расходятся, а сзади «сшиваются» с помощью ферментов. Цепь РНК постепенно растет и выдвигается из комплекса РНК-полимеразы.
Терминация процесса транскрипции происходит в участке-терминаторе, который распознается РНК-полимеразой благодаря спе-циальным белковым факторам терминации.

К 3′-концу синтезированной молекулы РНК присоединяется 150-200 адениновых нуклеотидов (поли-А), которые формируют «шлейф» необходимый для предотвращения ферментативного распада РНК. Еще ранее, когда был синтезирован 5′-конец, на нем был образован так называемый кэп (шапочка) — структура, представляющая собой метилированный остаток гуанозинтрифосфата, который защищает РНК от гидролиза.
В большинстве случаев в результате транскрипции не получается готовая РНК. «Сырая» РНК должна еще пройти процесс процес-синга, при котором происходят ее модификационные изменения и она становится функционально активной. Каждый тип РНК эукариот подвергается своим модификациям. Созревание РНК у эукариот осуществляется сложно. Во-первых, у эукариот сущест-вует ядро, которое отделено от цитоплазмы ядерной мембраной. В ядре осуществляется образование первичных транскриптов, которые имеют большую длину, чем цитоплазматическая мРНК, участвующая в трансляции. Следовательно, образованию зрелой мРНК у эукариот предшествует удале-ние интронов из последовательности гяРНК- транскрипта (этот процесс называется сплай-сингом от англ. to splice —сплетать, сращивать). После удаления последовательностей, соответствующих интронам, происходит соединение участков, которые транскрибированы с экзонов. Для точного узнавания в составе интронов есть специфические после-довательности — сигналы. В результате сплайсинга молекулярная масса мРНК уменьшается в 10 раз. Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосо-мам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция (от лат. translatio) -передача.
Перевод последовательности нуклеотидов в молекуле и-РНК в последовательность аминокислот в полипептидной цепи — молекуле белка.
Трансляция — синтез полипептидной цепи с использованием мРНК в роли матрицы. В трансляции участвуют три типа РНК: мРНК является информационной матрицей; тРНК доставляют аминокислоты и узнают кодоны; рРНК вместе с белками образуют рибосо-мы, которые удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи.
У прокариот рибосомы связываются с вновь синтезированной молекулой и-РНК сразу же после её отделения от ДНК. У эукариот и-РНК должна выйти из ядра в цитоплазму. Этот перенос и-РНК осуществляется специальными белками, которые образуют комплекс с молекулой РНК. Эти же белки защищают и-РНК от повреждающего действия цитоплазматических ферментов.
Синтез белка происходит в рибосомах. Рибосома-это немембранный органоид клетки, который состоит из двух субъединиц: боль-шой и малой. В функционирующей рибосоме имеется функциональный центр, который состоит из двух центров Аминоацильного (А— Центр узнавания аминокислоты) и Пептидильного (Р—Центр присоединения аминокислоты к пептидной цепочке). Строение рибосомы изучил коллектив, которым руководил российский учёный Александр Спирин. Сначала методом центрифугирования рибосома была выделена из клетки и рассмотрена под электронным микроскопом. С помощью биохимических манипуляций рибо-сому развернули, получили ленту. К ленте стали добавлять различные химические вещества, так выяснили, что в её состав входит рибосомная РНК и белок, которые чередуются в шахматном порядке. В цитоплазме на тот конец и-РНК с которого начинался синтез молекулы в ядре (3ʹ- конца) вступает малая субчастица рибосомы и начинает двигаться до старт-кодона (АУГ ). Далее к старт-кодону подходит т-РНК с комплементарным кодоном (соответствующим аминокислоте метионин), только после этого к малой субъединице рибосомы присоединяется большая субъединица и формируется функциональная рибосома. Первая т-РНК переносит аминокис-лоту метионин, которая оказывается в пептидильном участке рибосомы. Эта аминокислота будет присоединять к себе растущую цепь белка. Эта фаза трансляции называется инициация.
Параллельно с фазой инициации в цитоплазме происходит активизация аминокислот. Она происходит с помощью специальных фермен-тов аминоацил-т-РНК-синтетаз в присутствии АТФ. Аминокислоты присоединяются к акцепторному концу т-РНК в соответ-ствии с триплетом антикодона,
Фаза элонгации. Далее начинается фаза элонгации, во время которой происходит непосредственный синтез полипептида. Синтез поли-пептидной цепочки происходит в большой субчастице рибосомы. В аминоацильный центр рибосомы поступают т-РНК с ами-нокислотами и если антикодон т-РНК комплементарен триплету и-РНК, то она временно присоединяется к нему. Вновь поступив-шая аминокислота ока-зывается рядом с аминокислотой метионин и между карбоксильной группой метионина и аминогруппой другой Ак образуется пептидная связь, результатом чего является образование дипептида. т-РНК, которая принесла аминокислоту метионин, покидает рибосому, а т-РНК с другой аминокислотой перемещается в пептидильный центр, рибосома передвигается относительно и-РНК, и в рибосому поступает новая т-РНК со своей аминокислотой и в случае её комплементарности триплету в аминоацильном центре между этой аминокислотой и второй Ак вновь образуется пептидная связь, а пептид увеличится ещё на одну аминокислоту. Дальнейшее удлинение пептидной цепочки проис-ходит путём повторения предыдущих действий.
Таким образом, в стадии элонгации происходит последовательное наращивание полипептидной цепи по одной аминокислоте в соответствии с порядком кодонов в молекуле и-РНК.
Фаза терминации. Удлинение полипептидной цепи продолжается до тех пор, пока на пути рибосом не окажется один из термини-рующих (бессмысленных) триплетов и-РНК (УАА, УАГ, УГА). Этим триплетам не соответствует ни одна аминокислота. В области этих триплетов под действием ферментов происходит гидролитическое расщепление связи между пептидом и последней т-РНК. В ре-зультате освобождается готовый белок.

ДЛЯ ЗАКРЕПЛЕНИЯ ИЗУЧЕННОГО МАТЕРИАЛА ПЕРЕЙДИТЕ ПО ДАННОЙ ССЫЛКЕ: https://learningapps.org/view28216393 И ВЫПОЛНИТЕ ЗАДАНИЯ.
Выполните самостоятельно.
Задание 1. Ответьте на вопросы теста.
1. Материальным носителем наследственной информации является: А) иРНК Б) тРНК В) ДНК Г) хромосомы
2. В основе индивидуальности организмов лежит: А) строение белков организма Б) строение клеток В) функции клеток Г) строение аминокислот
3. ДНК клетки несет информацию о строении: А) белков, жиров, углеводов Б) белков и жиров В) аминокислот Г) белков
4. В одном гене закодирована информация: А) о структуре нескольких белков Б) о структуре одной из цепей ДНК В) о первичной структуре одной молекулы белка Г) о структуре аминокислоты
5. Какой из нуклеотидов не входит в состав ДНК: А) тимин Б) урацил В) гуанин Г) аденин
6. Какой из фактов подтверждает, что ДНК является генетическим материалом клетки: А) количество ДНК в клетках одного организма постоянно Б) ДНК состоит из нуклеотидов В) ДНК локализована в ядре клетки Г) ДНК представляет собой двойную спираль
7. В какой клетке человека нет ДНК: А) зрелый лейкоцит Б) зрелый эритроцит В) лимфоцит Г) нейрон
8. Транскрипцией называется: А) процесс образования иРНК Б) процесс удвоения ДНК В) процесс образования белковой молекулы Г) процесс соединения иРНК с аминокислотами
9. Синтез иРНК начинается с: А) разъединения молекулы ДНК на две нити Б) удвоения каждой нити В) взаимодействия РНК-полимеразы с геном Г) расщепления гена на нуклеотиды
10. Где синтезируется иРНК: А) в рибосомах Б) в цитоплазме В) в ядрышке Г) в ядре
11. Какую информацию содержит один триплет ДНК: А) о последовательности аминокислот в белке Б) об одном признаке организма В) об одной аминокислоте Г) о начале синтеза иРНК
12. Генетический код вырожден, потому что: А) одна аминокислота шифруется одним кодоном Б) несколько аминокислот шифруются одним кодоном В) между кодонами есть «знаки препинания» Г) одна аминокислота шифруется несколькими кодонами
13. Сколько комбинаций составляется из нуклеотидов: А) 61 Б) 64 В) 20
14. Трансляция – это: А) синтез полипептидной цепи на рибосомах Б) синтез тРНК В) синтез иРНК по матрице ДНК Г) синтез рРНК
15. Антикодоны тРНК комплементарны: А) кодонам рРНК Б) кодонам ДНК В) кодонам иРНК Г) всем указанным кодонам
16. Количество тРНК равно: А) количеству всех кодонов ДНК Б) количеству кодонов иРНК, шифрующих аминокислоты В) количеству генов тРНК Г) количеству белков в клетке
17. Второй этап биосинтеза заключается в: А) узнавании и присоединении аминокислоты к тРНК Б) снятии и «списывании» информации с ДНК В) отрыве аминокислоты от тРНК на рибосоме Г) создание иРНК
18. Синтез белка завершается в момент: А) появления на рибосоме «знака препинания» Б) истощения запасов ферментов В) узнавания кодона антикодоном Г) присоединения аминокислоты к тРНК
19. На полисоме синтезируется: А) одна молекула белка Б) несколько молекул различных белков В) несколько молекул одинаковых белков Г) возможны все варианты
20. Известно, что клетки многоклеточного организма имеют одинаковую генетическую информацию, но содержат разные белки. Какая из гипотез наиболее верна? А) разнообразие белков не зависит от особенностей клетки Б) в каждом типе клеток реализуется только часть генетической информации В) присутствие белков в клетке зависит не от генетической информации
21. Постройте последовательность реакций биосинтеза белка, выписав буквы в необходимом порядке: А) «списывание» информации с ДНК Б) узнавание антикодоном тРНК своего кодона на иРНК В) отщепление аминокислоты от тРНК Г) поступление иРНК на рибосомы Д) присоединение аминокислоты к белковой цепи с помощью фермента
22. Однозначность генетического кода проявляется в том, что каждый триплет кодирует: А) несколько аминокислот Б) не более двух аминокислот В) три аминокислоты Г) одну аминокислоту
23. Транспортная РНК – это: А) аминокислота Б) глюкоза В) липид Г) нуклеиновая кислота
24. Какой триплет тРНК комплементарен кодону ГЦУ на иРНК: А) ЦГТ Б) АГЦ В) ГЦТ Г) ЦГА
25. Сколько нуклеотидов в гене, кодирующем последовательность 60 аминокислот в молекуле белка: А) 60 Б) 120 В) 180 Г) 240
Задание 2. Выберите три правильных ответа.
1. Каковы особенности реакций биосинтеза белка в клетке? А) реакции носят матричный характер: белок синтезируется на иРНК Б) реакции происходят с освобождением энергии В) на химические реакции расходуется энергия АТФ Г) реакции сопровождаются синтезом АТФ Д) ускорение реакций осуществляется ферментами Е) синтез белка происходит на внутренней мембране митохондрий
2. В чем проявляется взаимосвязь биосинтеза белка и окисления органических веществ? А) в процессе окисления освобождается энергия, которая расходуется в ходе биосинтеза белка Б) в процессе биосинтеза образуются орган. вещества, которые используются в ходе окисления В) в процессе фотосинтеза используется энергия солнечного света Г) через плазматическую мембрану в клетку поступает вода Д) в процессе биосинтеза образуются ферменты, которые ускоряют реакции окисления Е) реакции биосинтеза белка происходят в рибосомах с выделением энергии
3. Какие из указанных процессов относятся к биосинтезу белка? А) Рибосома нанизывается на иРНК Б) в полостях и канальцах ЭПС накапливаются органические вещества В) тРНК присоединяют аминокислоты и доставляют их к рибосоме Г) перед делением клетки из каждой хромосомы образуются по две хроматиды Д) присоединенные к рибосоме две Ак взаимодействуют между собой с образованием пептидной связи Е) в ходе окисления органических веществ освобождается энергия
Задание 3. Установите соответствие.


Задание 4. Решите задачи.
1. Определите последовательность нуклеотидов на и-РНК, антикодоны т-РНК и аминокислотную последовательность соответствующего фрагмента молекулы белка (используя таблицу генетического кода), если фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ГТГ-ЦЦГ-ТЦА-ААА.
2. Одна из цепей ДНК имеет последовательность нуклеотидов: ЦАТ-ГГЦ-ТГТ–ТЦЦ–ГТЦ… Найдите иРНК, последовательность аминокислот. Объясните, как изменится структура молекулы белка, если произойдет удвоение четвертого триплета нуклеотидов в цепи ДНК?
3. В биосинтезе белка участвовали т-РНК с антикодонами: УУА, ГГЦ, ЦГЦ, АУУ, ЦГУ. Определите нуклеотидную последовательность участка каждой цепи молекулы ДНК, который несет информацию о синтезируемом полипептиде, и число нуклеотидов аденин, гуанин, тимин, цитозин в двухцепочечной молекуле ДНК.
Пройдите контрольный тест:
На вопросы теста можно ответить перейдя по ссылке: https://onlinetestpad.com/er74wt6xnpjse

0 Комментариев